Home



and big data to sustainably provide for our planet’s growing population.
Learn more about how our researchers are engineering for tomorrow.
and big data to sustainably provide for our planet’s growing population.
Learn more about how our researchers are engineering for tomorrow.





Extension Precision Ag Blog

Variable-Rate Fertilizer Application Considerations
It’s not hard to miss large fertilizer spreaders on the roads or in the fields at this time of the ...

Estimating Peanut Harvest Losses
Losses during peanut harvest are common and can be caused by improper setup, operation and/or performance of the harvest equipment ...

Pesticide Application Considerations with Spray Drones
With increased availability and interest in spray drones recently, we are starting to see more people getting into the spray ...
Precision Poultry Farming Blog

Tracking Poultry Locomotion with AI Technology
Poultry locomotion is an important indicator of animal health, welfare, and productivity. Traditional methodologies such as manual observation or the ...

2024 Georgia Precision Poultry Farming Conference
The UGA Poultry Science Department and Extension team will host the 2024 Georgia Precision Poultry Farming Conference virtually on May ...
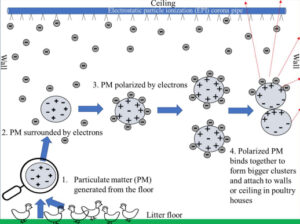
Electrostatic particle ionization technology for air quality management in poultry houses
In the USA, cage-free (CF) housing systems have gained popularity as an ethical and humane poultry farming method. However, the ...


